Это вторая часть из двух. В первой части больше теории, вторая часть более практическая.
Оглавление:
- Процессы в тканях при растяжке (ч.1)
1.1. Мышечная ткань (ч.1)
1.2. Соединительная ткань (ч.1)
1.3. Сосуды (ч.1)
1.4. Периферические нервы (ч.1) - Влияние растяжки на разные системы организма (ч.1)
2.1. Мышечная ткань (ч.1)
2.2. Соединительная ткань (ч.1)
2.3. Сосуды (ч.1)
2.4. Внутренние органы (ч.1)
2.5. Стрессовая адаптация и психическое состояние (ч.1) - Оптимальные методы развития гибкости
3.1. Нервная регуляция процесса растягивания
3.2. Недостатки баллистического и пассивного растягивания
3.3. Активное растягивание
3.4. Некоторые факторы, влияющие на развитие гибкости
3.5. Растягивание при травме - Использование знаний о растягивании при построении тренировочного занятия по хатха-йоге
4.1. Значение разминки для растягивания
4.2. Особенности рефлекторного эффекта растягивания
4.3. Особенности времени суток
4.4. Частота и длительность растягивания
4.5. Сочетание растягивания с другими видами тренинга - Приложения
5.1. Сравнение структуры коллагена и мышцы
5.2. Описание нервно-мышечных веретен
5.3. Описание нервно-сухожильных веретен
5.4. Характеристика системы суставных рецепторов
5.5. Анатомические половые различия
3. Оптимальные методы развития гибкости
Итак, основная приложение сил для обретения гибкости – соединительная ткань, окружающая мышечные волокна. Существуют разные методики, физиологический эффект которых мы и проанализируем.
Но для начала рассмотрим роль в процессе растягивания нервной регуляции.
3.1. Нервная регуляция процесса растягивания
Кроме собственно свойств мышечной и соединительной тканей, рассмотренных выше, в процессе растягивания существенную роль играет неврологический компонент.
Существуют внутримышечные, мышечно-сухожильные и суставные рецепторы. Подробнее о них см. в приложении. Все эти рецепторы сигнализируют об изменении длины мышцы и положения сустава, и способствуют управлению движением. Человек может повысить такую чувствительность, причем можно развить повышенное восприятие скорости движения, и слабо чувствовать изменение длины, либо наоборот. Заметим, что после обучения точного управления определенному движению процесс его корректировки (переобучение) потребует в 2 раза больше времени, чем процесс первичного усвоения.
Основными рецепторами, чьи сигналы влияют на процесс растягивания, будут мышечные (интрафузальные волокна). В ответ на процесс растягивания они запускают рефлекс растяжения. Он заставляет растянутую мышцу сокращаться, пытаться вернуться в исходное положение. Этот процесс имеет несколько особенностей:
- Чем выше сила растяжения, тем сильнее нервный стимул на сокращение.
- Сигнал от рецепторов мышечных веретен медленно адаптирующийся, т.е. – уровень дискомфорта от растянутой мышцы падает медленно.
- Прерывание адаптации на стимул сразу начинается заново.
- По достижении значительного растяжения, сопротивление движению внезапно снижается, несмотря на то, сигнал от рецепторов остается.
Рецепторы же мышечно-сухожильных веретен (Гольджи), вопреки принятому раньше мнению, реагируют только при очень сильном растягивании.
При регулярном (статическом) растягивании происходит адаптация к сигналам как местная (растянутая ткань даёт меньше импульсов в мозг), так и на уровне головного мозга. При этом кроме адаптации к сигналам от конкретного участка происходит и адаптация к процессу растягивании, как к таковому. В частности, человек обучается тянуть ткани более плавно, с меньшей силой, потому что так сигналы о дискомфорте значительно ниже.
Как будет описано ниже, при статическом растягивании рефлекс растяжения снижается, напряжение мышцы снижается, облегчая процесс расслабления.
Однако для обретения пластических изменений в мышце ПОЛНОЕ расслабление мышцы НЕ является необходимым.
Дело в том, что увеличение гибкости мышечной и соединительной тканей мышцы реализуется как раз за счет рефлекса растяжения. При растягивании и включении рефлекса растяжения сильное напряжение ложится на отдельные микро элементы тканей. В них происходят мелкие «травмы» – отслоение титина от миозина, частичное повреждение титина, уменьшение поперечных сшивок коллагена и пр. Эти «микротравмы» затем адекватно восстанавливаются.
3.2. Недостатки баллистического и пассивного растягивания
Баллистическое, или маховое растягивание в целом является негативным. Главным образом – за счет резких движений происходят травмы большего порядка, чем нужно. Травмированная ткань неадекватно замещается нефункционально ориентированной соединительной тканью, т.е. - рубцом. Также происходит неадекватная неврологическая адаптация – рецепторы стимулируются слишком сильно.
Пассивное растягивание выполняется без напряжения самого занимающегося, т.е. с помощью усилий партнера или специального аппарата. Выяснено, что оно зачастую слишком болезненно, и не способствует сохранению гибкости. Т.е. – оно вызывает скорее эластичные, а не пластичные изменения. Выяснено, что чем больше расхождение между активной и пассивной растяжкой (в сторону превалирования пассивной), тем выше вероятность травмы.
3.3. Активное растягивание
Активное растягивание подразумевает статическое или медленное динамическое растягивание с сознательным сокращением мышц антагонистов (противоположных растягиваемой), либо же ещё и с сознательным сокращением растягиваемой мышцы.
Рассмотрим статическое растягивание. Оно предусматривает растягивание мышцы до момента, когда дальнейшее её движение ограничивается собственным напряжением. В этом положении достигнутое растяжение поддерживается в течение определённого периода времени, при этом происходит расслабление и снижение напряжения.
Подобный феномен расслабления может быть объяснён следующим:
- Рецепторы растяжения мышцы (то есть нервно-мышечные веретена) десенсибилизируются и со временем адаптируются к растяжению. Таким образом, рефлекс растяжения снижается.
- Если напряжение мышцы вследствие растяжения достаточно большое, происходит активация нервно-сухожильных веретен и суставных рецепторов, что вызывает аутогенный рефлекс торможения. Мышца постепенно производит максимальные изометрические сокращения в течение 5-10 с. Это сокращение вынуждает нервно-сухожильные веретена выпускать импульсы, что инициирует аутогенное торможение и расслабление мотонейрона растянутой мышцы.
- Мышца и соединительная ткань обладают зависимыми от времени механическими свойствами. Другими словами, при воздействии постоянной силы возникает постепенное изменение длины, наряду с плавным снижением напряжения.
- Кратковременное статическое растяжение снижает электрическую активность. Статическое растягивание приводит к незначительному снижению возбудимости двигательного пула.
- Фактическая длина мышцы меняется слабо, т.е. нагрузка растяжением будет изометрической. Доказано, что она вызывает значительно меньший болевой эффект, чем эксцентрическая нагрузка (пассивное увеличение длинны мышцы). Растягивание с практически не замечаемым болевым эффектом позитивно влияет на психическое состояние, тогда как боль может вызывать «подсознательное» стремление к травме.
По достижении момента первичного ограничения напряжением в активном растягивании используют либо изометрическое, либо изотоническое сокращение мышцы-антагониста (или группы антагонистичных мышц). В результате реципрокной иннервации должно снизиться напряжение в растягиваемой мышце. Именно благодаря этому явлению так же значительно снижается болевое ощущение при растягивании.
Выяснено, что при активной растяжке достаточным временем фиксации будет 30с. В случаях жёстких мышц, или неэффективного управления (за счёт неопытности) – до 60с.
Поскольку при растягивании нагрузка ложится на отдельные микро-элементы тканей, есть смысл за тренировку делать несколько «подходов» на растягивание нужного участка. Либо же делать многократные возвратно-поступательные подходы.
Что касается медленных динамичных движений, они эффективны для увеличения пластичности мышцы в случае наличия статической фиксаций в конечной точке хотя бы на 10-15 секунд.
При статическом и медленном динамическом растягивании постепенно улучшается проведение нервно-мышечной передачи импульсов, что делает растягивание в дальнейшем более эффективным.
Активная гибкость имеет более высокую степень корреляции с уровнем спортивных достижений (г = 0,81), чем пассивная (г = 0,69). А в сочетании с силовыми упражнениями корреляция достигает (0,91).
Это просто объясняется тем, что чем сильнее мышца-антагонист, тем большее усилие она может применить по отношению к растягиваемой мышце. Однако нужно заметить, что применение слишком большой силы не нужно, т.к. это вызывает чересчур сильное сокращение растягиваемой мышцы и недостаточное её удлинение для улучшения пластичности.
В любом случае, силовые занятия нужны для укрепления ткани, чтобы не перейти предел её эластичности. Как описывалось выше, гибкость также помогает силовым возможностям мышцы.
Повышенная пластичность мышцы, достигнутая активным растягиванием, является самой стабильной. Часто для поддержания достигнутой гибкости достаточно нескольких, или даже одного интенсивного занятия в месяц.
3.4. Некоторые факторы, влияющие на развитие гибкости
Гибкость не является общей, а является специализированной для различных суставов. Так, в разном возрасте в детстве и юности соотношение гибкости разных суставов меняется.
Соматотип не коррелирует с гибкостью.
Установлено, что женщины обладают большей гибкостью, чем мужчины. Различия в уровнях гибкости между мужчинами и женщинами могут быть обусловлены рядом факторов, включая анатомические и физиологические. Подробнее см. приложение 5.5.
У жесткой мышцы рано наступает сопротивление движению – на последних 40% (у нормальной – на последних 20%).
При многократных растягивающих движениях у жесткой мышцы на последних подходах рефлекс начинается позже. У нормальной мышцы нет.
У жесткой мышцы в результате растяжки упруговязкий компонент снижается, а неврологический нет.
3.5. Растягивание при травме
Воспаление представляет собой сосудистую и клеточную реакцию, направленную на восстановление поврежденных тканей. Степень повреждения ткани зависит от величины силы, воздействующей на волокна. Степень воспаления, в свою очередь, прямо пропорциональна величине или степени повреждения. Воспалительная реакция представляет собой динамичную и непрерывную последовательность хорошо согласующихся явлений. 5 основных признаков воспаления: покраснение, припухлость, повышение температуры, боль и изменение или потеря функции.
Заключительными этапами воспалительного процесса являются восстановление (ремонт) и ремоделирование. Восстановление связано с пролиферацией капилляров и фибробластов (клеток, синтезирующих коллагеновые волокна). Поэтому фазу восстановления нередко называют фиброплазией. В этот период времени новые коллагеновые волокна произвольно ориентируются и хорошо растворимы; установленные связи еще очень хрупкие. Во время ремоделирования коллагеновый синтез продолжается, наряду с реориентацией коллагеновых фибрилл в направлении нагрузки и образованием нормальных поперечных связей между фибриллами. Таким образом, микроскопически архитектура коллагеновых волокон изменяется в сторону более организованной структуры. При относительно значительном повреждении наблюдаются обширные рубцы. Сила и пластические характеристики рубцовой ткани зависят от образования и плотности межмолекулярных ковалентных связей, а также от ориентации отдельных коллагеновых волокон.
Рубцевание, особенно обширное, представляет собой определенную проблему. Мышца очень медленно восстанавливает свою силу, сухожилия же, вследствие менее интенсивного кровоснабжения, еще медленнее.
Сила — не единственный важный физический параметр, на который влияют рубцы. В результате образования рубцовой ткани утрачивается эластичность. За счёт этого растяжения имеют тенденцию рецидивировать (повторяться). Чем чаще возникают растяжения в определённом участке мышцы, тем больше количество рубцовой ткани и тем выше вероятность рецидива травм.
Итак, по мере занятий коллагеновые сшивки могут уменьшаться в рубце. Рубцы ремоделируются медленно.
Рубец соединительной ткани должен привыкнуть, а уже потом можно тянуться по новой – менять ориентацию коллагена и повышать предел прочности на разрыв.
Вывод: в зависимости от тяжести травмы от 10 до 30 дней желательно вообще прекратить или свести к минимуму растягивание травмированного участка.
Затем возвращаться к растягиванию этого участка постепенно. Дискомфорт, возникающий при ремоделировании ткани нормален. Но нужно быть осторожным, при реабилитации мышца может порваться еще до боли.
4. Использование знаний о растягивании при построении тренировочного занятия по хатха-йоге
4.1. Значение разминки для растягивания
Для гибкости важно повышение температуры соединительной и мышечной ткани вследствие разминки. Увеличение температуры приводит к снижению предела прочности соединительной ткани, вследствие чего можно было бы ожидать увеличение количества разрывов. Однако с повышением температуры увеличивается степень растяжимости, и по факту, разминка на самом деле предотвращает вероятность разрывов и других травм.
Кроме этого, повышается эффективность реципрокной иннервации, что позволяет мышцам-антагонистам быстрее и эффективнее сокращаться и расслабляться.
Предварительное повышение тонуса симпатоадреналовой системы и/или уровня СО2 в крови в результате определённых разминочных упражнений повысит температуру.
Но с точки зрения развития гибкости разминка не обязательно должна иметь большую продолжительность и выраженный подъем симпатоадреналового тонуса. Более того, она может быть не общей, а локальной для каждой мышцы. Например - возвратно-поступающие движения на растягивание с постепенным увеличением амплитуды движения.
4.2. Особенности рефлекторного эффекта растягивания
Уже обсуждалось слияние процесса растягивания на внутренние органы через моторно-висцеральные и кожно-висцеральные рефлексы. Для «общей» тренировки желательна стимуляция большого количества рефлекторных зон (мест повышенной концентрации рецепторов).
Наиболее эффективными для воздействия на внутренние органы являются статическая или многократная динамическая стимуляция, с помощью медленных динамических движений с небольшой фиксацией (5-15с) в конечной фазе движения. Рекомендуемое количество повторений – от 10. Слишком быстрые движения или малое количество повторений при динамическом режиме будет менее эффективным.
Растягивание тканей грудной клетки и плечевого пояса через стимуляцию симпатических узлов вызовет повышение общего тонуса (вегетативного и ЦНС), что хорошо вначале тренировки. Растягивание таза и ног через стимуляцию парасимпатического крестцового сплетение вызовет некоторое общее торможение, что хорошо в конце тренировки.
С другой стороны, динамическое растягивание как таковое за счет частых раздражений разных участков нервной системы вызовет общую активацию. А статическое растягивание с постепенной адаптацией и понижением как раздражающего сигнала, так и ответа на него – общее торможение. Поэтому вначале тренировки и растягивание ног и таза будет к месту, но в относительно динамичном режиме. Большую часть мышц и связок ног, таза, и значительную часть спины эффективно растягивать стоя на ногах, т.к. ноги при этом являются сильными рычагами.
При статическом растягивании за несколько лет наступает выраженная адаптация общего неврологического ответа, что является одним из факторов стрессовой адаптаций. С другой стороны, несколько теряется эффект «раздражения растягиванием». Такое «раздражение» оказывается возможным за счет динамического режима. В случае использования динамического режима, он должен применяться в первой части занятия, как дающий общий активирующий эффект.
Любое растягивание, даже медленное и применимое к «тормозящему» парасимпатическому сплетению, оказывает раздражающее (стрессовое) воздействие на определенные регуляторные системы. Поэтому по окончании выполнения упражнений на растягивание желательно выполнить упражнение на успокоение.
Исключение составляют люди с повышенным парасимпатическим тонусом, для которых тренировка должна заканчиваться состоянием активации.
4.3. Особенности времени суток
Существенное снижение роста происходит в первые часы после пробуждения, 38% - в первые 1,5 часа. Восстанавливается рост на 70% - в первые ночные часы. Такие перепады роста объясняются увеличением толщины межпозвоночных дисков, за счет набора воды в первые часы постоянного горизонтального положения. Причины такого набора до конца не ясны, диуретик, принятый в ночные часы, не меняет картины.
Из-за увеличенного объема межпозвоночных дисков упражнения на позвоночник могут быть опасными для него в первые полтора часа после пробуждения.
4.4. Частота и длительность растягивания
Мы выяснили, что безопасных и эффективных способов растягивания два – статический и медленный динамический с небольшими фиксациями в конце.
При статическом для гибкости достаточно 60с фиксации, но для реализации рефлекторного воздействия на внутренние органы желательна фиксация около 3 минут. Замети, что в основном это касается растягивания тканей корпуса.
При динамическом растягивании желательно около 5-10 повторений, с фиксацией в конечных положениях 5-15с. Либо такое же количество возвратно-поступательных движений, как правило, выполняемых с увеличением амплитуды движения.
Оптимальная частота занятий на растягивание в неделю для улучшения гибкости 2-4 раза. Чаще - эффективность, как правило, повышаться не будет. На частоту влияет индивидуальные особенности соединительной ткани («жесткость на вытяжение»). Опытному практику, с хорошим контролем расслабления/напряжения мышц при условии нежесткой ткани часто достаточно 2 раз в неделю интенсивного растягивания. Людям с «жесткой» соединительной тканью желательно не меньше 3 раз. Максимум 4–5, нужно давать тканям отдых.
Двухразовые занятия могут быть избыточным стрессом для определенных систем организма, и скорее будут нежелательными.
4.5. Сочетание растягивания с другими видами тренинга
Мы уже выяснили, что занятия на гибкость способствуют силе, и наоборот.
Т.к. динамика активирует, а статическое растягивание «тормозит» общую активность, то растягивание в динамическом режиме следует практиковать в первой части занятия, до силового тренинга. К тому же блок динамического растягивания вызовет повышение эндорфинов и кортизола. Кортизол является гормоном «разрушения», и его повышение как раз будет к месту пред силовой нагрузкой для улучшения внутриклеточных регуляторных сигналов мышечной ткани, запускающих адаптивные процессы. Эндорфины повышают способность психической компоненты по проведению силового тренинга, а также оказывают на мышцу анаболический эффект.
Статическое растягивание эффективно выполнять в конце тренинга, и после каждого силового упражнения или группы упражнений, направленных на одну группу мышц. Это улучшит кровообращение в мышце сразу по окончании силового упражнения.
Одного «успокоительного растягивания» недостаточно для полноценного успокоения, поэтому в конце тренировки в любом случае необходимо выполнить «шавасану» (длительное расслабление лежа). В этом случае «финальное растягивание» может быть недолгим (около 5 минут).
Что касается совмещения растягивания и других видов физической активности, то его можно использовать и до и после. До – для повышения гибкости во время «активности». После – в статическом виде для некоторого успокоения, хотя можно пользоваться и другими методами успокоения (дыхательные, шавасана).
5. Приложения
5.1. Сравнение структуры коллагена и мышцы

5.2. Описание нервно-мышечных веретен
Основными тензорецепторами (рецепторами давления) мышцы являются нервно-мышечные веретена. Поскольку мышечные волокна заключены в веретенообразную капсулу (оболочку соединительной ткани, окутывающую рецептор), их называют интрафузальными волокнами. Экстрафузальные волокна являются обычными сократительными единицами мышцы. Веретена прикрепляются к обоим концам экстрафузальных волокон и, таким образом, располагаются параллельно им. Следовательно, при растягивании мышцы веретено также подвергается растягиванию.
Существует два основных типа интрафузальных волокон: волокна ядерного мешочка и волокно ядерной цепочки. Эта несократительная структура находится в центральном, или экваториальном, участке интрафузального волокна. Отсюда и название — интрафузальное волокно ядерного мешочка. На дистальных, или полярных, концах волокна ядерного мешочка находятся исчерченные сократительные филаменты, которые возле веретен прикрепляются к экстрафузальным волокнам.
Второй тип интрафузального мышечного волокна — волокно ядерной цепочки — более тонкое и короткое, чем волокно ядерного мешочка. Оно содержит всего один ряд ядер, расположенных в виде цепочки по несократительному экваториальному участку. Подобно волокнам ядерного мешочка, полярные концы волокна ядерной цепочки также состоят из исчерченных сократительных филаментов. Их концы нередко соединяются с волокнами ядерного мешочка, которые, в свою очередь, прикрепляются к эндомизию экстрафузальных волокон.
Чувствительные нервы веретен
Существует два вида чувствительных нервных (афферентных) окончаний в каждом веретене: первичные и вторичные. Первичные окончания заканчиваются, спиралевидно окружая центральный участок волокна ядерного мешочка и выпуская боковое ответвление к волокну ядерной цепочки. Афферентные аксоны первичных нервных окончаний относятся к волокнам большой группы I. Чтобы отличить эти чувствительные нервные окончания от других, относящихся к этой же группе, большие афференты веретена называют афферентами группы Iа.
Первичные нервные окончания имеют очень низкий порог растяжения и поэтому легко возбуждаются. Их реакции могут быть как фазными (динамическими), так и тоническими. Фазная реакция определяет интенсивность или скорость растягивания путем изменения частоты нейронных импульсов во время растягивания. Частота разрядки быстро увеличивается при начальном растягивании. Затем, по достижении новой длины, она снижается до постоянного уровня, соответствующего новой тонической длине. Следовательно, тоническая реакция определяет длину мышцы. Иными словами, первичные нервные окончания определяют длину и скорость растягивания.
Вторичные нервные окончания образуют разветвленные, напоминающие цветок окончания. Они содержатся в основном в околоэкваториальном сегменте (вблизи экватора) волокон ядерной цепочки. Аксоны вторичных нервных окончаний относятся к афферентным волокнам группы II. В отличие от первичных нервных окончаний, вторичные нервные окончания определяют только тоническую длину мышцы.
Эти мотонейроны вызывают сокращение мышечных филаментов в полярных концах интрафузальных мышечных волокон. При сокращении этих концов в центральном экваториальном участке наблюдается пассивное растягивание (там находятся чувствительные нейронные рецепторы). Таким образом, активация гамма-мотонейронов ЦНС может увеличивать количество растяжения, испытываемого чувствительными нервными окончаниями.
Двигательные нервы веретена
Двигательные нервы, иннервирующие каждое интрафузальное мышечное волокно в его полярных участках, называются у-эффереитными волокнами, образующими фузимоторную систему. Различают два вида у-аксонов в зависимости от их воздействия на первичные и вторичные чувствительные нервные окончания. Стимулирование так называемого статического у-аксона (ys) может усиливать чувствительность первичного нервного окончания к длине, не влияя (или незначительно влияя) на восприятие (чувствительность) скорости. С другой стороны, динамические у-аксоны (yd) могут существенно усиливать восприятие скорости первичного нервного окончания, практически не влияя на чувствительность к длине.
Функция у-системы заключается в контроле чувствительности веретена к растягиванию. Этот процесс сенситизации веретена при помощи у-эфферентов известен как у-смещение. Активация у-мотонейронов приводит к сокращению или укорачиванию интрафузальных мышечных волокон в их полярных участках. При сокращении интрафузальных волокон экваториальный участок растягивается. Эта центральная деформация приводит к перетягиванию колец в аннулоспиральную часть первичных окончаний и увеличению интенсивности разрядки афферентов групп Iа и II.
Вторая функция у-мотонейронов — поддержание чувствительности веретена во время сокращений мышцы. Когда мышцы укорачиваются, веретено также пассивно укорачивается. Это пассивное сближение двух концов веретена снимает напряжение как с его первичных окончаний (разгрузка веретена), так и со вторичных; такая разгрузка лишает головной мозг информации из веретена относительно изменений мышечной длины. Чтобы предотвратить разгрузку веретена и отрегулировать его чувствительность, у-мотонейроны активируют.
Процесс возбуждения нервно-мышечного веретена
Процесс возбуждения нервно-мышечного веретена можно охарактеризовать следующим образом. Во-первых, к нервно-мышечному веретену прикладывают минимальный растягивающий стимул. Во-вторых, в чувствительном нейронном окончании происходит изменение степени проницаемости, что приводит к производству генерирующего тока (передача заряда через мембрану нервного терминала). Последний, в свою очередь, вызывает деполяризацию, которая называется генератор потенциала. При несколько большей степени растягивания его амплитуда в окончаниях нервно-мышечного веретена увеличивается. Когда деполяризация достигает порога, возникает проведённый потенциал действия. При дальнейшем увеличении степени растягивания может наблюдаться серия проводимых нервных импульсов.
5.3. Описание нервно-сухожильных веретен
Нервно-сухожильные веретена — это чувствительные к сокращению механорецепторы скелетных мышц млекопитающих, иннервируемые быстропроводящими афферентными нервными волокнами, имеющими большой диаметр.
Нервно-сухожильные веретена находятся почти исключительно у апоневрозов или мышечно-сухожильных соединений, и довольно мало в самих сухожилиях. Местонахождение нервно-сухожильных веретен обеспечивает их экстремальную чувствительность к любому изменению напряжения отдельных мышечных волокон, к которым они прикреплены.
Нервно-сухожильные веретена располагаются последовательно, в отличие от нервно-мышечных веретен, расположенных параллельно мышечным волокнам. В исследовании Пенга количество мышечных волокон, соединенных последовательно с нервно-сухожильными веретенами, колеблется от 3 до 50. Каждое нервно-сухожильное веретено, как правило, иннервируется отдельным быстропроводящим афферентным нервным волокном.
Нервно-сухожильные веретена контролируют все степени мышечного напряжения. В то же время они более всего чувствительны к напряжению, производимому в результате мышечных сокращений. Предполагают также, что нервно-сухожильные веретена вносят свой вклад в сознательные восприятия. Это предположение основано на том факте, что импульс из нервно-сухожильных веретен достигает коры головного мозга, той его части, которая осуществляет интерпретацию сенсорной активности тела.
По мере увеличения напряжения в группе мышечных волокон нервно-сухожильные веретена и их афференты посылают увеличивающееся число сигналов в ЦНС. Они проявляют очень низкий порог и весьма ощутимую динамическую чувствительность, и способны сообщать об очень незначительных изменениях. Эти чувствительные нервы заканчиваются в спинном мозгу на небольших интернейронах, которые затем ингибируют клеточные тела мотонейронов, активирующих сокращающуюся мышцу. Данный процесс называется автогенным ингибированием, так как сокращения мышцы ингибируются её собственными рецепторами. Результирующее снижение мышечной силы уменьшает активацию нервно-сухожильных веретен и количество ингибиторной обратной связи, поступающей в ЦНС из этих мышц. Такое мгновенное ингибирование обратной связи способствует увеличению напряжения в мышцах.
За прошедшее время появилось немало ошибочных представлений о нервно-сухожильных веретенах. Одно из них касается взаимосвязи между нервно-сухожильными веретенами и измерением растягивающих усилий. Нервно-сухожильные веретена нередко называют «рецепторами растяжения». Этот термин подразумевает, что пассивное напряжение, сопровождающее растягивание мышцы, может также представлять собой адекватный стимул для нервно-сухожильных веретен. Нервно-сухожильные веретена, действительно, могут активироваться в результате пассивного напряжения, однако их порог для данного вида стимулов является очень высоким. Поэтому для активизации нервно-сухожильных веретен необходимо очень интенсивное растягивание. Кроме того, разрядка нервно-сухожильных веретен очень редко продолжается во время поддерживаемого растяжения мышцы.
5.4. Характеристика системы суставных рецепторов
|
Морфология |
Местонахождение |
Источник нервных волокон |
Бихевиоральиая характеристика |
|
|
I |
Шарообразные частицы (100 мкм • 40 мкм). покрытые тонкой оболочкой в трехмерных кластерах, состоящих из 3-8 частиц |
Фиброзные капсулы суставов (в поверхностных слоях) |
Небольшой миелинированный (6-9 мкм) |
Статические и динамические механорецепторы: низкий порог. Медленноадаптирующиеся |
|
II |
Конические частицы (280 мкм • 120 мкм),покрытые толстой оболочкой, отдельные или в кластерах по 2-4 частицы |
Фиброзные капсулы суставов (в глубоких субсиновиальных слоях). Суставные жировые тела |
Средний миелинированный (9-12 мкм) |
Динамические механорецепторы: низкий порог, быстроадаптирующиеся |
|
III |
Веретенообразные части цы (600 мкм • 100 мкм), покрытые тонкой оболочкой, отдельные или в кластерах но 2-3 |
Поверхность суставных связок (коллатеральные и внутренние) |
Большой миелиннрованный (13-17 мкм) |
Динамические механорецепторы: высокий порог. медленноадаптирующиеся |
|
IV |
(а) Трехмерные сплетения нервных волокон |
Фиброзные капсулы суставов. Суставные жировые тела. Адвентициальные оболочки кровеносных сосудов |
Очень маленькие миелинированные (2-5 мкм) и немнелинированные (< 2 мкм) |
Болевые рецепторы: очень высокий порог, неадаптирующиеся. Гемочувствительные (к аномальным метаболитам ткани) болевые рецепторы |
|
IV |
(б) Свободные немиелинированные нервные окончания |
Суставные связки (коллатеральные и внутренние) |
5.5. Анатомические половые различия
Одним из анатомических факторов, обеспечивающих женщинам большую степень гибкости, является различие между участками таза мужчин и женщин. Кости таза мужчин, как правило, более тяжелые и крупные, граница входа в таз не закруглена, полость менее широкая; саркоседалищная вырезка, лонная дуга и крестец более узкие, а вертлужная впадина более компактная, чем у женщин. А так как у женщин более широкие бедра, диапазон движения в тазовой области у них больше.
В то же время и у женщин наблюдаются различия в структуре таза, которые влияют на диапазон движения. Чаще всего используется классификация видов таза, предложенная Колдуэллом и Молоем (1933), основанная на форме верхнего края таза.
На рисунке приведено схематическое изображение четырех типов таза и влияние характерных изменений структуры таза на механизмы движения:
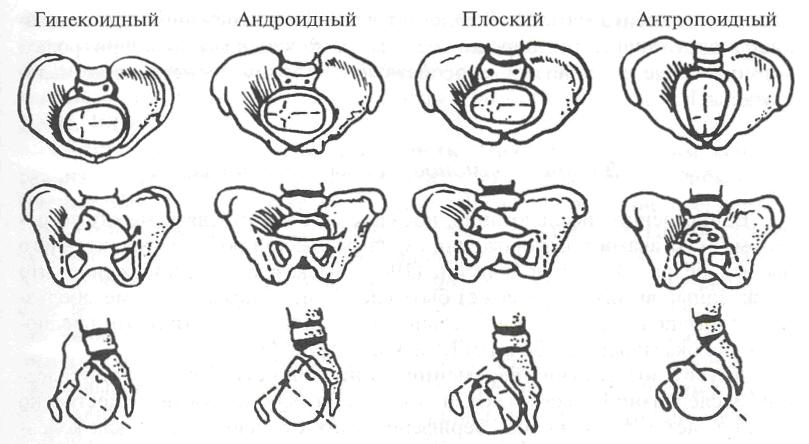
Гинекоидный, или женский, тип таза встречается чаще всего — почти у 50% женщин. Он характеризуется округленным или слегка овальным входом; лобковая дуга образует угол, равный приблизительно 90°.
Андроидный, или воронкообразный, тип таза встречается примерно у 20% женщин. Он характеризуется сердцеобразным верхним краем, клиноподобным входом и углом лобковой дуги 60-75°. Женщины, имеющие этот тип таза, сталкиваются с определенными трудностями при родах.
Плоский тип таза наименее часто встречается у мужчин и женщин. Лишь у 5% обследованных был выявлен этот тип таза. Он характеризуется равномерным сужением переднезаднего диаметра и смещением крестца вперед между подвздошными костями.
Примерно 20% женщин обладают антропоидным типом таза. Он характеризуется удлиненным переднезадним размером и укороченным поперечным диаметром. Обычно таз этого типа настолько большой, что роды протекают довольно легко.
Высказывается предположение, что девочки имеют более высокий потенциал развития гибкости после пубертатного периода и особенно амплитуды сгибания туловища вследствие более низко расположенного центра тяжести и меньшей длины ног по сравнению с мальчиками.
Кроме того, для женщин также характерна большая амплитуда выпрямления локтевого сустава. Это объясняется более короткой верхней кривизной олекранального отростка локтевого сустава у женщин по сравнению с мужчинами.